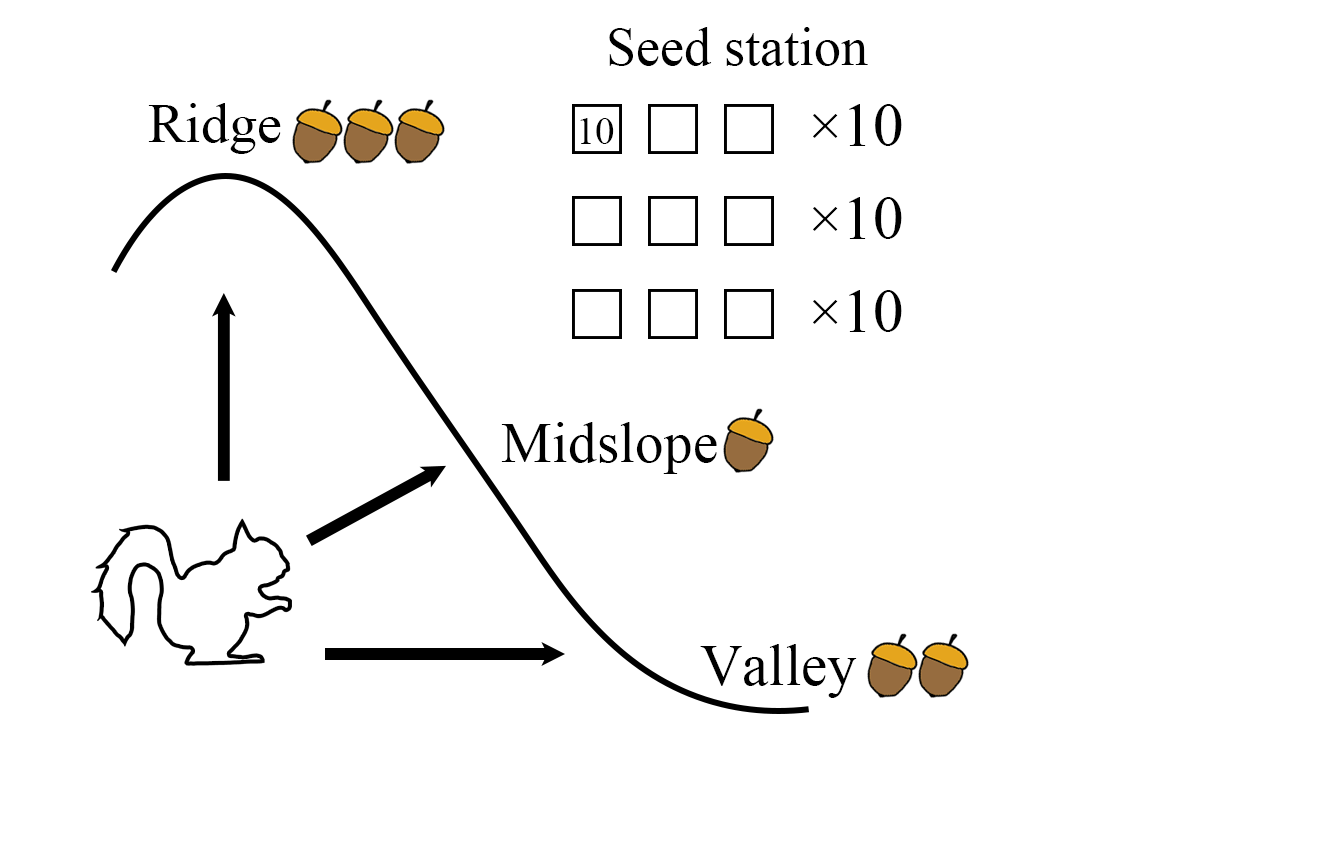
IntroductionDispersal limitation is the failure of a seed to reach all regeneration sites suitable for its germination after leaving the parent. The failure reasons include seed factors (i.e., seed dispersal mode, dispersal ability, seed size, and yield) and environmental factors (i.e., wind, water flow, and topography) (Han and Wang, 2002; Thomson, 2011). Previous studies have shown that dispersal limitation played an important role in Changbai Mountain forests for the Jaccard coefficient of seed rain and neighboring mature trees decreased sharply with increasing neighborhood radius (Li et al. , 2012) and in the habitat fragmentation of tropical forest, where species diversity is rich and seeds are mainly dispersed by animals (Cordeiro et al. , 2009). Considering niche differentiation and dispersal limitation simultaneously may better explain successional mechanisms in tropical forests (Dalling et al. , 2002), and dispersal limitation may be more important than ecological niche differentiation in determining species composition in habitats where dispersers are scarce and forest cover is low (Dent and Estrada-Villegas, 2021). For example, dispersal limitation due to the disappearance of seed dispersers such as birds and mammals in tropical forests has been a major factor in the inability of later successional species to spread into young forests (Abbas et al. , 2021). Therefore, birds and mammals are believed to play an important role in seed dispersal (Willson, 1993; Glyphis et al. , 1981). Previous studies suggested that mammals and birds were probably the main dispersers of palm seeds (Zona & Henderson, 1989), and mainly focused on the effects of large mammals and birds on seed dispersal (Gosper et al. , 2005). Seeds from excrement or seeds dropped during consumption by fruit-feeding primates could survive and germinate (Corlett & Lucas, 1990), and these seedlings had a higher survival rate (Pearson & Theimer, 2003). The composition and dynamics of large mammal and bird communities have undergone significant alterations, primarily attributable to pervasive overhunting and extensive habitat loss on a global scale (Carpenter et al. , 2020; Fernández-Palacios et al. , 2021). Furthermore, this has resulted in a significant reduction in the population of these large herbivores, leading to extinctions in certain cases, as well as a conspicuous reduction in the average body weight of large mammalian species. Evidenced by a shift from an average of approximately 14 kilograms at the 95th percentile to a mere 4 kilograms (Wenny & Levey, 1998), this trend underscores a substantial change in mammalian community structure. Consequently, this transformation has elevated the ecological significance of smaller fauna, particularly rodent species, in the process of seed dispersal.While feeding on plant seeds and fruits, rodents carry some seeds and fruits to other places for storage to cope with food shortages (Zhanget al.et al. , 2015), and to help seeds escape predators, ultimately leading to successful seed dispersal and regeneration (Hirschet al. , 2012). Therefore, dispersal hoarders, mainly rodents, are prominent seed dispersers of plants (Roth & Vander Wall, 2005). These rodents can spread viable seeds to different favorable environments (Jansen & Forget, 2001), some seeds buried can be retrieved by animals for food, while others would germinate successfully and achieve seedling settlement to a large extent. This animal storage behavior, especially decentralized storage, is profitable in reducing the competition for resources and space between seedlings and mother trees, to promote seed germination and seedling formation and to realize plant regeneration (Xiao & Zhang, 2016; Yang et al. , 2020). Food storage animals usually have an advantage over general foragers in finding storage food (Stapanian & Smith, 1978), this may be related to not only the difficulty of retrieval and the hidden conditions (Tamura et al. , 1999), but also the directional dispersal preference of certain storage animals. For example, scatter-hoarding rodents would transport seeds to high-density microsites (such as the tree canopy) and redistribute them for storage (Dimitri & Longland, 2017), resulting in the seed survival rate and the dispersal pattern, and ultimately affecting the regeneration pattern of vegetation (Yu et al. , 2017). The repeated movement of seeds and the distance to the habitat as the two main factors determining seed dispersal in rodents (Perea, 2011), and chemosensory information from predators and parasitoids also influence foraging decisions of storage rodents, and thus seed dispersal (Sunyeret al. , 2013).Seed dispersal and seedling establishment are the key stages to the mountain forest succession and regeneration, and their failure would limit the recolonization of farmland (Standish et al. , 2007). Some studies exploring seed dispersal constraints are currently receiving increasing attention from naturalists, especially topography-specific structures (Muscarella et al. , 2020; Wanget al. , 2014; Freitas et al. , 2010). The topography in mountainous and hilly areas plays a significant role in regulating the distribution of the solar radiation and precipitation, this can effectively reveal the microclimatic state of the local environment and showcases variations in the soil depth and nutrient content across different locations (McDonald et al. , 1996). As an important topographical factor, slope position directly affects vegetation through geomorphic processes and controls the spatial redistribution of resource factors through morphological changes, thus indirectly affecting the distribution of vegetation (Kikuchi, 2001; Yang, 2005). Furthermore, slope position also has an effect on vegetation cover and its ability to recover after disturbance, with lower slopes having higher vegetation cover and height, while upper slopes are more susceptible to disturbances such as forest fire and deforestation (Pereira et al. , 2016). The higher the slope of the mountain, the more spatially isolated it is and the more susceptible it is to fire and logging disturbances (Han et al. ,2018). Furthermore, crop yields are generally lower in the upper slope than those in the lower slope due to differences in soil types (McPhee, 1997), and acorns on the upper slope are more susceptible to insects than those on the lower slope (Yu, 2003). Scatter hoarders also weigh the pros and cons of cache difficulty against the risk of theft, with higher risk of predation often implying higher seed establishment rates (Muñoz & Bonal, 2011; Steele et al. , 2014). Compared to the hilltop area, there are usually many exposed rocks and vegetation providing safe shelter for rodents in midslope (Meliyo et al. , 2014), this may lead to better seed storage and seedling regeneration. Treeline recruitment was seriously affected by the high rate of seed theft by rodents (Pansing et al. , 2007). Therefore, the dispersal limitation is more pronounced on the upper slope than on the lower slope (Han et al. ,2018). However, the relationship between food storage rodents and their dispersal plants often changes depending on environmental conditions, with mutual benefits and seed predation occurring (Vander Wall, 2010), and there is no strict consistency.As a result, differences in vegetation regeneration exist between different slope positions, yet the reasons for these disparities remain unknown. Could rodent-mediated seed dispersal serve as a critical factor in explaining the regeneration variations among slope positions? To quantitatively measure variations in seed dispersal across slope positions, the current study explored rodent abundance and tracked seed removal rate, seed fate, and seed dispersal distance by establishing experimental plots at different slope positions of the Q. wutaishanica -rodent ecosystem in the Dongling Mountains situated in the North of Taihang Mountains. Thus, addressing the following questions: 1) Do slope positions significantly influence rodent-mediated seed dispersal? 2) What about the seed dispersal efficiency among different slope positions? 3) Are there definite seed dispersal limitations among slope positions? Based on the aforementioned inquiries, we formulate the following preliminary hypotheses: 1) Concerning seed dispersal, in accordance with the Mid-domain Effect (MDE) Model (Colwell and Lees, 2000), the dispersal rate is expected to exhibit a unimodal pattern as slope position increases, characterized by an initial rise followed by a subsequent decline. 2) In terms of faunal and floral abundance, according to the Resource Availability Hypothesis (Coley, 1985), with increasing slope position, an equilibrium point of resource utilization is anticipated to exist, resulting in a pattern of initial abundance escalation followed by a subsequent decline.Materials and MethodsStudy regionThe Beijing Forest Ecosystem Research Station (40°00’-40°03’N, 115°26’-115°30’ E) was located in Dongling Mountains, the northern part of the Taihang Mountains. The region belongs to warm temperate semi-humid continental monsoon climate, and its average annual temperature is 5~10℃ and mean annual precipitation is 500~600 mm, mainly precipitate from July to August. The main soil type is brown soil and its depth is about 50 cm. The zonal vegetation is warm temperate deciduous broad-leaved and coniferous mixed forest, especially the Q. wutaishanica forest, which is widely distributed and plays a significant role in the conservation of water and soil, the maintenance of biodiversity and the regulation of climate in the Taihang Mountains. To minimize heterogeneity among forest types dominated by Q. wutaishanica , ten transects were established from the foot to the top of every mountain’s western slope (i.e., each transect occupied a different elevation segment of the slopes), forming a single elevational gradient of montane forests (1020 m-1770 m) (Xu et al. , 2017). According to Parker’s (1982) classification method widely used in previous studies (Coppoletta et al. , 2016; Guarín et al. , 2005; Clinton et al. , 1993) and the topographic characteristics of Dongling Mountains, the transects were divided into three slope positions: ridge, midslope, and valley, forming three continuous sampling lines of oak forest at the different slope positions.Seed markingThe seed of Q. wutaishanica was collected during the mature period in 2020, and stored in the refrigerator at 4 ℃ for the germination rate of the oak seeds stored for seven days only slightly decreased and there was no significant difference compared to fresh seeds (Chen and Yan, 2019). Therefore, fresh and healthy seeds were selected within 48 hours by the water flotation and visual inspection method. The seed marking was completed by referring to the original method proposed by Zhang and Wang (2001) and made some necessary changes. A micro-drill was used to drill a small hole with a diameter of 0.3 mm at the bottom of the seed to avoid damaging the core. Small consecutively numbered plastic labels were made to track the seed fate, and 10 cm thick wires were used to pass through the holes in the seeds to connect the plastic labels with the seeds. Tags are generally exposed to the surface of the ground in most cases, so there is a high chance of finding them after the seeds have been buried by rodents. According to Kempter’s (2018) research, labels have some influence on seed dispersal, but would not change with slope position and other factors concerned.Seed releaseThe seeds were released in 10 transects set along the elevation gradient in the Q. wutaishanicaforest in the Taihang Mountains from August 26 to October 14 in 2020. According to the elevation gradient, each transect line was classified into three slope positions: ridge (the upper 1/3 of each transect), midslope (the middle 1/3 of each transect), and valley (the lower 1/3 of each transect). In total, there were 90 seed stations, 10 transects×3 slope positions×3 seed stations, and there were 10 seeds released at each 1 m × 1 m seed station. For repeated experiments, the scale of concern was the community and the factor of interest was the slope position, with 30 seed stations set for each slope position. Considering that seed fates changed little after 15 days, observations were made only once a week. Therefore, it is observed two or three times a week in the early stage and only once a week after 15 days, in order to assess the harvest of the seeds and their removal by small rodents (Fig.1). Seed fates were classified as six types: (1) intact in situ (IIS), (2) eaten in situ (EIS), (3) eaten after removal (EAR), (4) intact after removal (IAR), (5) scatter hoarded (SH) or (6) missing (M). We recorded the seed code numbers, measured the distance between the tagged seed and its original seed station where the seed was removed, and marked the specific cache locations. During March and May of the following year, we investigated and identified the seedlings established from seeds scattered by rodents according to the tagged plastic labels.Rodent composition structureTrapping plots were set at about 500 m from the seed dispersal experiment region at each transect and its three slope positions in the Taihang Mountains to minimize interference. Three live traps, formed a trapping plot, were placed at each slope position in August 2019 and 2020, respectively. Rodents were captured using 40 cm× 25 cm× 20 cm live stell wire traps with peanuts as bait. The trapping experiment lasted one week per transect and slope position, and the trapping period was one month per year. From August to October 2020, an infrared camera was set up in each seed dispersal area of each slope position to monitor rodent activity.Seedling surveysWe selected 10 transects to survey seedlings in the Taihang Mountains from October 3 to October 6 in 2019 before seed release experiment. Three slope positions (ridge, midslope and valley) were selected for each transect to investigate the abundance of Q. wutaishanicaseedlings. The seedling survey experiment was repeated three times and a total of 90 (3 slope positions × 10 sample strips × 3 replicates) seedling data were obtained.Data analysisAll statistical analysis were performed in R version 4.1.1 (R Core Team, 2021). The seed removal time of three slope positions was compared using Cox Regression Analysis, and seed fate and dispersal distance were analysed using seed tracking experiments. Differences in seed fate (i.e., IIS, EIS, EAR, IAR, SH and M) among three slope positions and the influence of the distance group on the removal distance were examined using a Generalized Linear Model (GLM). Tukey’s HSD post hoc test was applied for multiple comparisons of seed fates, seed removal rate, and seed dispersal distance among three slope positions. The number of seedlings was analysed using One-way analysis of variance (ANOVA) with post hoc tests using the Least Significant Difference (LSD)method.ResultsRodent species composition and abundanceThere were 38, 32 and 3 captured rodents in valley, midslope, and ridge, respectively, including Apodemus peninsulae , Apodemus agrarius , Sciurotamias davidianus , Tamiops swinhoei , andSciurus vulgaris (Table 1). A. peninsulae was the most common, accounting for 64.4% of the total, followed by S. davidianus and T.s swinhoei , and their percentages were 17.8% and 12.3%. Specifically, the first four rodent species mentioned above were captured in midslope and their relative abundances in the midslope were 68.6%, 3.1%, 9.4% and 18.8%, respectively. All five species of rodents were captured in valley, and A. peninsulae and S. davidianus were common, accounting for 63.2% and 26.3%, respectively. Only three rodents were captured in ridge, including A. peninsulae , A. agrarius , and T. swinhoei .Seed removal rateMost of the seeds released in valley and midslope, except the ridge, were carried by rodents within 11 days (Fig.2). Cox regression analysis showed that seed removal rates were significantly higher in midslope (Z=10.69, P <0.001) and valley (Z=10.21,P <0.001) than that in ridge. Furthermore, the seed removal rate was significantly higher in midslope (Z=2.24P =0.0252) than in valley. The seed removal rates in midslope and valley almost had no change after 11 days and then became basically stable after 22 days.Seed fateAlmost all seeds were kept intact (ISS) in ridge, while half of the seeds at valley and midslope were eaten by rodents (EAR, EIS) (Fig.3). The ratios of IIS, EIS, EAR, IAR and M in valley (IIS: t =14.112,df =87, P <0.001; EIS: t =6.504,df =87, P <0.001; EAR: t =8.619,df =87, P <0.001; IAR: t =2.707,df =87, P =0.0220; M: t =8.800, df =87,P <0.001) and midslope (IIS: t =13.363,df =87, P <0.0001; EIS: t =5.488,df =87, P <0.001; EAR: t =9.328,df =87, P <0.0001; IAR: t =4.410,df =87, P =0.0001; M: t =4.470, df =87,P =0.0001) differed significantly from those in ridge. The proportion of SH in ridge (t =1.892, df =87,P =0.1470) and midslope (t = 3.478, df = 87,P = 0.0023) had not significant difference with that in valley, while there was significant difference in the proportion of SH between ridge and midslope (t =3.379, df =87, P =0.0031).Seed dispersal distanceThe seeds dispersed by the rodent in ridge were all within 3 m and were mainly concentrated in 1-3 m (Fig.4). The maximum dispersal distance of midslope was almost 15 m, with a major concentration around 5 m. The average seed dispersal distance was significantly higher in midslope (t =3.449, df =207,P =0.002) than that in ridge, indicating that the seed was dispersed farther in midslope. In contrast, the dispersal distance of the seeds in ridge was finite.Seedling surveyThe mean value of seedling was 1.43 cm as a whole, while those in ridge, midslope and valley were 1.13 cm, 1.83 cm and 1.33 cm. The variance test statistic showed that the number of seedling in different slope positions were significantly different (F=4.567, P = 0.013<0.05). The number of seedling was significantly less in ridge than those in midslope (P =0.004, SE=0.239) and valley (P =0.039, SE=0.239). There were no significant differences for the number of seedling between midslope and valley (P =0.404, SE=0.239).DiscussionSeedling regenerationAccording to our investigation, the number of seedlings in ridge was lower than midslope and valley, this is consistent with the results of Takahashi et al. (2010) and O’Brien & Escudero (2022), supporting our hypothesis (2) as mentioned above. The reason is that soil in ridge is shallower and drier than other slope position, creating droughty and frost-prone conditions, which can be detrimental to seedling germination and early establishment (Fenner & Thompson, 2005; Takahashi et al. , 2010). Furthermore, as the ridge slope increased near the top, airflow velocity increased, increasing snow flux and snow transport, leading to an increase in snow accumulation (Toloui-Semnani & Johnson, 2019) and intensifying transpiration of pioneer seedlings (Renison et al. , 2015). This further exacerbates the challenges faced by seedlings trying to establish themselves in ridge. Seedlings at ridge showed wilting and needle discoloration due to low photosynthetic carbon gain, reduced soil water potential, low symbiotic infection of seedlings with mycorrhizal fungi, limited root growth, and ultimately seedling dehydration, which may explain the higher than ninety percent mortality rate of seedlings in the ridge region (Smith et al. , 2009). As long as the processes of freezing, tumbling, and burial persist at ridge, it will be very difficult to establish seedlings in these areas (Butler et al. , 2009). The ecosystem stability of mid-elevation zones at the altitude between 500 and 2000 m were particularly stable, while high-elevation zones (above 2000 m) and low-elevation zones (below 500 m) were comparatively vulnerable (Geng et al., 2019). This is, the communities of middle region being more stable compared to the other two regions (Jin et al., 2022). Furthermore, the forest litter layer may act as a barrier, making it difficult for seeds to contact the soil, thus affecting tree regeneration in the Taihang Mountains (Yang et al., 2014). These natural challenges make ridge areas particularly inhospitable for seedling regeneration and survival. Therefore, topographic fragmentation, special climate and soil conditions lead to poor vegetation regeneration in Taihang Mountains, with simple layers and low coverage, especially at the ridge of the mountain.Rodent abundanceAmong the five recorded rodent species, the Apodemus peninsulaewas the most common in the Taihang Mountains, and the midslope and valley had more rodents than ridge. According to the ’landscape of fear’ theory, vegetated habitats can provide shelter and improve survival chances for animals, so animals prefer vegetated habitats over open high-risk habitats (Laundré et al. , 2001), small rodents were found in greater numbers in forested sparse grassland areas (Afonso, 2021), and nests around shrubs on the slopes could avoid the negative effects of rainfall and uphill runoff on burrows (Jiang et al. , 2017). Furthermore, plants generally exhibit better growth in midslope compared to ridge and valley (Liao et al. , 2021; Ohsawa & Ide, 2008), and the vertical structure of vegetation distribution on the mountain exhibits a central peak pattern (Lomolino, 2001). A similar feature was also found that the distribution of alpha diversity of plant community in the Taihang Mountains had unimodal distribution on the elevational gradient, which was in line with the theory of ”diversity peaked at the intermediate altitude” (Liang, 2022). Therefore, the midslope and valley may have more rodents than the ridge. Significantly, the distribution of small mammals exhibits a mid-elevation peak, i.e., a single-peaked distribution at mid-elevation (McCain, 2010) according to the MDE (Colwell and Lees, 2000), meaning that the midslope should have more rodents than ridge and valley. Unfortunately, this study showed that there were more rodents in the valley, which is inconsistent with hypothesis (2), but there is little difference in rodent abundance between the valley and the midslope. This may be due to the limitation of plant growth by forest water sources as vegetation is primarily driven by energy supply (Roebroek et al. , 2020). According to the energy limitation hypothesis (Colwell and Lees, 2000), light, water and other conditions in mid-slope may be moderate, but not as abundant as in valley, some rodent species may adapt to this medium energy limitation environment, but other species may not adapt well, reducing their diversity and quantity. Our results of rodent abundance were obtained based on camera trap monitoring and traditional survey methods, the applicability of the high resolution, the large breadth, and the high sensitivity of infrared camera may allow better access to animal behavioral data and may improve the results in future.Seed removal rate and seed fateThe seed removal rate and the seed collected proportion by rodents was lowest in ridges, which is consistent with our hypothesis (1). On the contrary, we found that the midslope had the highest seed removal rate and proportion collected. Seed removal rate is a critical metric for understanding the dynamics of seed dispersal and regeneration within an ecosystem, reflecting a composite of various ecological activities, including those of seed predators, primary seed dispersers, and secondary seed dispersers (Hambuckers et al. , 2020; Milotić et al. , 2018). So, this rate is not merely a quantitative measure but also provides insights into the qualitative aspects of seed dispersal mechanisms and their effectiveness under different environmental conditions (Carlo & Morales, 2016). Seeds in ridges exhibit limited dispersal, encountering challenges in germination primarily attributed to restricted movement towards suitable germination sites (Forget, 1990). The limitation in seed dispersal is chiefly due to the diminished or absent role of key seed dispersers, particularly rodents and birds. According to the niche limitation hypothesis (Romanuk & Kolasa, 2002), the scarcity of resources and lower temperatures at high elevations would increase the intensity of competition so that only rodent species adapted to these conditions and survived in these regions. Ecological niches at high altitudes may only be suitable for some specific rodent species. Previous studies highlight the importance of specific seed dispersers for large-seeded plants is underscored (Moreira et al. , 2017). For example, the critical function of various bird sizes in seed dispersal (Godínez-Alvarez et al. , 2020), the interspecific interactions of rodents for enhancing seed dispersal effectiveness in Japanese walnut (Okawa et al. , 2023), and the dual reciprocal roles in pollination and seed dispersal of dispersal-storage rodents (Xiao, 2021). These studies emphasize the necessary contribution of vertebrates in overcoming the challenges of seed dispersal, confirming the integrated part of these animals in maintaining ecosystem balance and plant reproduction (La Mantia et al. , 2019). Thus, the limiting effects of the absence of dispersers on seed dispersal are evident, which not only hinders the movement of seeds to suitable germination sites, but may also has long-term effects on the balance of the ecosystem as a whole.An interesting observation is that rodent-mediated seed dispersal was more active in midslope, instead of the valley and ridge. Within our study, the EIS of Q. wutaishanica on the mid-slope was found to be lower than the average EIS rate (74%) reported in previous studies (Li and Zhang, 2001). Midslope’s higher seed removal rate and scatter-hoarding rate imply greater opportunities for seed dispersal and establishment. This is inconsistent with the idea that seed release at the top of the mountain was an order of magnitude greater than at the bottom of the mountain (Katul, 2012). This is mainly because: 1) the seeds are likely to roll downhill when they break away from the mother tree, which can lead to superimposed seed shadows from different mother trees (Ohsawa et al. , 2007; Schupp et al. , 2019) on midslope and valley. Furthermore, some valleys may accumulate seeds at the bottom or in valley areas, as they may be natural meeting points for wind or water. (Boland, 2017). 2) Rodents prefer to spread seeds horizontally or downward because it consumes less energy (Li and Zhang, 2003), but the absence of rodents may limit seed dispersal and seedling regeneration in ridge. Although birds are capable of dispersing and establishing seeds in high mountainous or other high-altitude areas (Watanabe, 1994), Quercus species are only moderately preferred as food by birds (Wright, 2022). Consequently, the impact of birds on the dispersal of Quercus seeds is quite limited.Seed dispersal distanceThe dispersal distance reflects rodent foraging strategies and dispersing effectiveness to some extent (Wang and Yan, 2017). We found that the dispersal distances of the seeds were longer in midslope and valley than those in ridge where the dispersal distance in ridge was extremely limit (Fig.4). This phenomenon further validates the reasonableness of our hypothesis (1). Although seeds would tend to accumulate on lower slopes due to gravity and topography, and then valley had a higher seed density (Yamase and Sekioka, 2006), rodent-mediated seed dispersal is a central driver of horizontal seed dispersal as it requires less energy (Wang et al. , 2018), while in the vertical direction is just the opposite. Compared to the ridge and valley, the better vegetation and less disturbance on midslope attract more rodents to feed, and provide more diverse habitats for rodents to meet their requirements of spreading over a longer distance while avoiding predation (Bergstedt and Milberg, 2001). Furthermore, long-distance dispersal increases reproduction rates by allowing seeds to escape the high density of conspecifics in the vicinity of the mother tree, reducing competition and predators, and providing them with the opportunity to find suitable colonization sites (Novaes et al. , 2020) according to the negative density-dependent hypothesis (Metzet al. , 2010; Jansen et al. , 2014). In our study, the dispersal distance was concentrated within 15 m, which is consistent with previous studies (Chen et al. , 2022). The dispersal distance was mostly around and within 5 m, and the seed removal rate exhibited an initial rapid decline followed by a gradual slowdown, indicating that the rodents accelerated the possession of food, which might be caused by competition (Jenkins and Peters, 1992).These results, as mentioned above, were helpful in understanding the dispersal of seeds mediated by rodents in the Q. wutaishanicapopulation at different slope positions. Further research still needs to interpret the seed dispersal and vegetation regeneration pattern at a larger temporal and spatial scale (Nathan and Muller-Landau, 2000). For example, Q. wutaishanica interannual variation and its subsequent effects on vegetation structure might have been overlooked (Wang and Smith, 2002).